
Adaptive immunity
The adaptive immune system is composed of highly specialized, systemic cells and processes that eliminate or prevent pathogenic challenges. Thought to have arisen in the first jawed vertebrates, the adaptive or "specific" immune system is activated by the “non-specific” and evolutionarily older innate immune system (which is the major system of host defense against pathogens in nearly all other living things). The adaptive immune response provides the vertebrate immune system with the ability to recognize and remember specific pathogens (to generate immunity), and to mount stronger attacks each time the pathogen is encountered. It is adaptive immunity because the body's immune system prepares itself for future challenges.
The system is highly adaptable because of somatic hypermutation (a process of accelerated somatic mutations), and V(D)J recombination (an irreversible genetic recombination of antigen receptor gene segments). This mechanism allows a small number of genes to generate a vast number of different antigen receptors, which are then uniquely expressed on each individual lymphocyte. Because the gene rearrangement leads to an irreversible change in the DNA of each cell, all of the progeny (offspring) of that cell will then inherit genes encoding the same receptor specificity, including the Memory B cells and Memory T cells that are the keys to long-lived specific immunity. Immune network theory is a theory of how the adaptive immune system works, that is based on interactions between the variable regions of the receptors of T cells, B cells and of molecules made by T cells and B cells that have variable regions.
A scanning electron microscope (SEM) image of a single human lymphocyte.
Functions
Adaptive immunity is triggered in vertebrates when a pathogen evades the innate immune system and generates a threshold level of antigen.The major functions of the adaptive immune system include:
the recognition of specific “non-self” antigens in the presence of “self”, during the process of antigen presentation.
the generation of responses that are tailored to maximally eliminate specific pathogens or pathogen infected cells.
the development of immunological memory, in which each pathogen is “remembered” by a signature antibody.
These memory cells can be called upon to quickly eliminate a pathogen should subsequent infections occur.
Effector cells
The cells of the adaptive immune system are a type of leukocyte, called a lymphocyte. B cells and T cells are the major types of lymphocytes. The human body has about 2 trillion lymphocytes, constituting 20-40% of white blood cells (WBCs); their total mass is about the same as the brain or liver. The peripheral blood contains 20–50% of circulating lymphocytes; the rest move within the lymphatic system.
B cells and T cells are derived from the same pluripotential hematopoietic stem cells, and are indistinguishable from one another until after they are activated. B cells play a large role in the humoral immune response, whereas T-cells are intimately involved in cell-mediated immune responses. However, in nearly all other vertebrates, B cells (and T-cells) are produced by stem cells in the bone marrow. T-cells travel to and develop in the thymus, from which they derive their name. In humans, approximately 1-2% of the lymphocyte pool recirculates each hour to optimize the opportunities for antigen-specific lymphocytes to find their specific antigen within the secondary lymphoid tissues.

In an adult animal, the peripheral lymphoid organs contain a mixture of B and T cells in at least three stages of differentiation:
naive cells that have matured, left the bone marrow or thymus, have entered the lymphatic system, but that have yet to encounter their cognate antigen,
effector cells that have been activated by their cognate antigen, and are actively involved in eliminating a pathogen and,
memory cells – the long-lived survivors of past infections.
Antigen presentation
Adaptive immunity relies on the capacity of immune cells to distinguish between the body's own cells and unwanted invaders.The host's cells express "self" antigens. These antigens are different from those on the surface of bacteria ("non-self" antigens) or on the surface of virally infected host cells (“missing-self”). The adaptive response is triggered by recognizing non-self and missing-self antigens.
With the exception of non-nucleated cells (including erythrocytes), all cells are capable of presenting antigen and of activating the adaptive response. Some cells are specially equipped to present antigen, and to prime naive T cells. Dendritic cells and B-cells (and to a lesser extent macrophages) are equipped with special immunostimulatory receptors that allow for enhanced activation of T cells, and are termed professional antigen presenting cells (APC).

Association between TCR and MHC class I or MHC class II
Several T cells subgroups can be activated by professional APCs, and each type of T cell is specially equipped to deal with each unique toxin or bacterial and viral pathogen. The type of T cell activated, and the type of response generated depends, in part, on the context in which the APC first encountered the antigen.
Exogenous antigens
Dendritic cells engulf exogenous pathogens, such as bacteria, parasites or toxins in the tissues and then migrate, via chemotactic signals, to the T cell enriched lymph nodes. During migration, dendritic cells undergo a process of maturation in which they lose most of their ability to engulf other pathogens and develop an ability to communicate with T-cells. The dendritic cell uses enzymes to chop the pathogen into smaller pieces, called antigens. In the lymph node, the dendritic cell will display these "non-self" antigens on its surface by coupling them to a "self"-receptor called the Major histocompatibility complex, or MHC (also known in humans as Human leukocyte antigen (HLA)).This MHC:antigen complex is recognized by T-cells passing through the lymph node. Exogenous antigens are usually displayed on MHC class II molecules, which activate CD4+ helper T-cells.
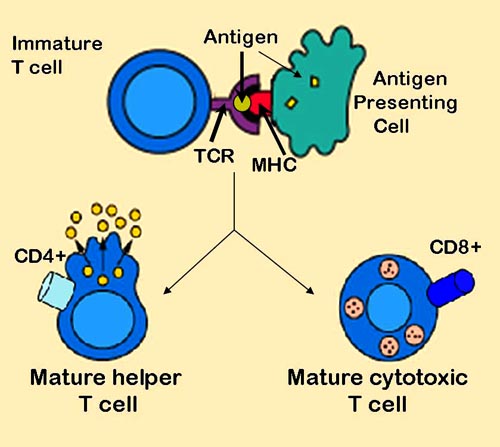
Antigen presentation stimulates T cells to become either "cytotoxic" CD8+ cells or "helper" CD4+ cells
Endogenous antigens
Endogenous antigens are produced by viruses replicating within a host cell. The host cell uses enzymes to digest virally associated proteins, and displays these pieces on its surface to T-cells by coupling them to MHC. Endogenous antigens are typically displayed on MHC class I molecules, and activate CD8+ cytotoxic T-cells. With the exception of non-nucleated cells (including erythrocytes), MHC class I is expressed by all host cells.
T lymphocytes
CD8+ T lymphocytes and cytotoxicity

Cytotoxic T cells (also known as TC, killer T cell, or cytotoxic T-lymphocyte (CTL)) are a sub-group of T cells which induce the death of cells that are infected with viruses (and other pathogens), or are otherwise damaged or dysfunctional.

Killer T cells—also called cytotoxic T lymphocytes or CTL-directly attack other cells carrying certain foreign or abnormal molecules on their surfaces
Naive cytotoxic T cells are activated when their T-cell receptor (TCR) strongly interacts with a peptide-bound MHC class I molecule. This affinity depends on the type and orientation of the antigen/MHC complex, and is what keeps the CTL and infected cell bound together.Once activated, the CTL undergoes a process called clonal expansion in which it gains functionality, and divides rapidly, to produce an army of “armed”-effector cells. Activated CTL will then travel throughout the body in search of cells bearing that unique MHC Class I + peptide.
When exposed to these infected or dysfunctional somatic cells, effector CTL release perforin and granulysin: cytotoxins which form pores in the target cell's plasma membrane, allowing ions and water to flow into the infected cell, and causing it to burst or lyse. CTL release granzyme, a serine protease that enters cells via pores to induce apoptosis (cell death). To limit extensive tissue damage during an infection, CTL activation is tightly controlled and generally requires a very strong MHC/antigen activation signal, or additional activation signals provided by "helper" T-cells (see below).
Upon resolution of the infection, most of the effector cells will die and be cleared away by phagocytes, but a few of these cells will be retained as memory cells. Upon a later encounter with the same antigen, these memory cells quickly
differentiate into effector cells, dramatically shortening the time required to mount an effective response.
Helper T-cells
CD4+ lymphocytes, or helper T cells, are immune response mediators, and play an important role in establishing and maximizing the capabilities of the adaptive immune response. These cells have no cytotoxic or phagocytic activity; and cannot kill infected cells or clear pathogens, but, in essence "manage" the immune response, by directing other cells to perform these tasks.
Helper T cells express T-cell receptors (TCR) that recognize antigen bound to Class II MHC molecules. The activation of a naive helper T-cell causes it to release cytokines, which influences the activity of many cell types, including the APC that activated it. Helper T-cells require a much milder activation stimulus than cytotoxic T-cells. Helper T-cells can provide extra signals that "help" activate cytotoxic cells.
Th1 and Th2: helper T cell responses

T helper activation
Two types of effector CD4+ T helper cell responses can be induced by a professional APC, designated Th1 and Th2, each designed to eliminate different types of pathogens. The factors that dictate whether an infection will trigger a Th1 or Th2 type response are not fully understood, but the response generated does play an important role in the clearance of different pathogens.
The Th1 response is characterized by the production of Interferon-gamma, which activates the bactericidal activities of macrophages, and induces B-cells to make opsonizing (coating) antibodies, and leads to "cell-mediated immunity" The Th2 response is characterized by the release of Interleukin 4, which results in the activation of B-cells to make neutralizing (killing) antibodies, leading to "humoral immunity". Generally, Th1 responses are more effective against intracellular pathogens (viruses and bacteria that are inside host cells), while Th2 responses are more effective against extracellular bacteria, parasites and toxins. Like cytotoxic T-cells, most of the CD4+ helper cells will die upon resolution of infection, with a few remaining as CD4+ memory cells.
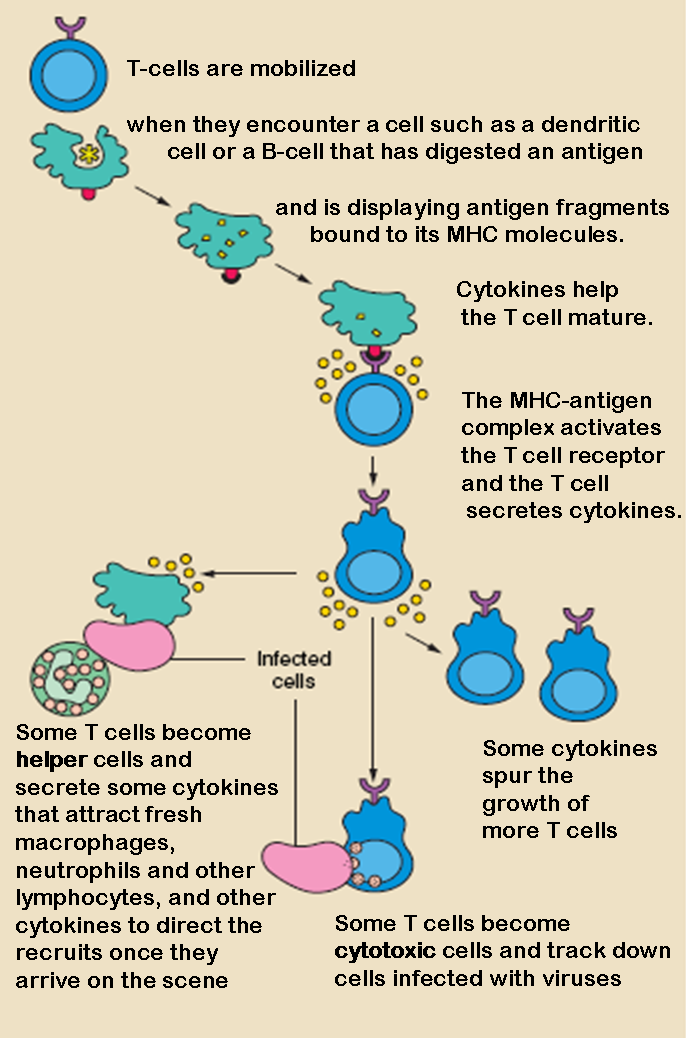
The T lymphocyte activation pathway. T cells contribute to immune defenses in two major ways: some direct and regulate immune responses; others directly attack infected or cancerous cells
HIV is able to subvert the immune system by attacking the CD4+ T cells, precisely the cells that could drive the destruction of the virus, but also the cells that drive immunity against all other pathogens encountered during an organisms' lifetime.
A third type of T lymphocyte, the regulatory T cells (Treg), limits and suppresses the immune system, and may control aberrant immune responses to self-antigens; an important mechanism in controlling the development of autoimmune diseases.T cells never act as antigen-presenting cells.
gamma/delta T cells

Representation of a gamma-delta T cell receptor
(gamma delta cells) possess an alternative T cell receptor (TCR) as opposed to CD4+ and CD8+ αβ T cells and share characteristics of helper T cells, cytotoxic T cells and natural killer cells. Like other 'unconventional' T cell subsets bearing invariant TCRs, such as CD1d-restricted Natural Killer T cells, γδ T cells exhibit characteristics that place them at the border between innate and adaptive immunity. On one hand, γδ T cells may be considered a component of adaptive immunity in that they rearrange TCR genes via V(D)J recombination, which also produces junctional diversity, and develop a memory phenotype. On the other hand however, the various subsets may also be considered part of the innate immune system where a restricted TCR and/or NK receptors may be used as a pattern recognition receptor. For example, according to this paradigm, large numbers of Vγ9/Vδ2 T cells respond within hours to common molecules produced by microbes, and highly restricted intraepithelial Vδ1 T cells will respond to stressed epithelial cells.
B lymphocytes and antibody production
B Cells are the major cells involved in the creation of antibodies that circulate in blood plasma and lymph, known as humoral immunity. Antibodies (or immunoglobulin, Ig), are large Y-shaped proteins used by the immune system to identify and neutralize foreign objects. In mammals there are five types of antibody: IgA, IgD, IgE, IgG, and IgM, differing in biological properties, each has evolved to handle different kinds of antigens. Upon activation, B cells produce antibodies, each of which recognizes a unique antigen, and neutralize specific pathogens.
Like the T cell receptor, B cells express a unique B cell receptor (BCR), in this case, an immobilized antibody molecule. The BCR recognizes and binds to only one particular antigen. A critical difference between B cells and T cells is how each cell "sees" an antigen. T cells recognize their cognate antigen in a processed form - as a peptide in the context of an MHC molecule, while B cells recognize antigens in their native form. Once a B cell encounters its cognate (or specific) antigen (and receives additional signals from a helper T cell (predominately Th2 type)), it further differentiates into an effector cell, known as a plasma cell.

The B lymphocyte activation pathway. B cells function to protect the host by producing antibodies that identify and neutralize foreign objects like bacteria and viruses.
Plasma cells are short lived cells (2-3 days) which secrete antibodies. These antibodies bind to antigens, making them easier targets for phagocytes, and trigger the complement cascade. About 10% of plasma cells will survive to become long-lived antigen specific memory B cells. Already primed to produce specific antibodies, these cells can be called upon to respond quickly if the same pathogen re-infects the host; while the host experiences few, if any, symptoms.

Structure 3D d'une immunoglobuline IgG

IgM pentamer structure

IgE structure

IgD Structure

Ig A structure
In other words

Alternative adaptive immune system
Although the classical molecules of the adaptive immune system (e.g. antibodies and T cell receptors) exist only in jawed vertebrates, a distinct lymphocyte-derived molecule has been discovered in primitive jawless vertebrates, such as the lamprey and hagfish. These animals possess a large array of molecules called variable lymphocyte receptors (VLRs for short) that, like the antigen receptors of jawed vertebrates, are produced from only a small number (one or two) of genes. These molecules are believed to bind pathogenic antigens in a similar way to antibodies, and with the same degree of specificity.
Immunological memory
When B cells and T cells are activated some will become memory cells. Throughout the lifetime of an animal these memory cells form a database of effective B and T lymphocytes. Upon interaction with a previously encountered antigen, the appropriate memory cells are selected and activated. In this manner, the second and subsequent exposures to an antigen produce a stronger and faster immune response. This is "adaptive" because the body's immune system prepares itself for future challenges. Immunological memory can either be in the form of passive short-term memory or active long-term memory.
Passive memory
Passive memory is usually short-term, lasting between a few days and several months. Newborn infants have had no prior exposure to microbes and are particularly vulnerable to infection. Several layers of passive protection are provided by the mother. In utero, maternal IgG is transported directly across the placenta, so that at birth, human babies have high levels of antibodies, with the same range of antigen specificities as their mother. Breast milk contains antibodies that are transferred to the gut of the infant, protecting against bacterial infections, until the newborn can synthesize its own antibodies.
This is passive immunity because the fetus does not actually make any memory cells or antibodies, it only borrows them. Short-term passive immunity can also be transferred artificially from one individual to another via antibody-rich serum.
Active Memory
Active immunity is generally long-term and can be acquired by infection followed by B cells and T cells activation, or artificially acquired by vaccines, in a process called immunization.
Immunization
Historically, infectious disease has been the leading cause of death in the human population. Over the last century, two important factors have been developed to combat their spread; sanitation and immunization.Immunization (commonly referred to as vaccination) is the deliberate induction of an immune response, and represents the single most effective manipulation of the immune system that scientists have developed.Immunizations are successful because they utilize the immune system's natural specificity as well as its inducibility.

The principle behind immunization is to introduce an antigen, derived from a disease causing organism, that stimulates the immune system to develop protective immunity against that organism, but which does not itself cause the pathogenic effects of that organism. An antigen (short for antibody generator), is defined as any substance that binds to a specific antibody and elicits an adaptive immune response.
Most viral vaccines are based on live attenuated viruses, while many bacterial vaccines are based on acellular components of micro-organisms, including harmless toxin components.Many antigens derived from acellular vaccines do not strongly induce an adaptive response, and most bacterial vaccines require the addition of adjuvants that activate the antigen presenting cells of the innate immune system to enhance immunogenicity.
Immunological diversity
Most large molecules, including virtually all proteins and many polysaccharides, can serve as antigens. The parts of an antigen that interact with an antibody molecule or a lymphocyte receptor, are called epitopes. Most antigens contain a variety of epitopes and can stimulate the production of antibodies, specific T cell responses, or both

An antibody is made up of two heavy chains and two light chains. The unique variable region allows an antibody to recognize its matching antigen.
A very small proportion (less than 0.01%) of the total lymphocytes are able to bind to a particular antigen, which suggests that only a few cells will respond to each antigen.
For the adaptive response to "remember" and eliminate a large number of pathogens the immune system must be able to distinguish between many different antigens,and the receptors that recognize antigens must be produced in a huge variety of configurations, essentially one receptor (at least) for each different pathogen that might ever be encountered. Even in the absence of antigen stimulation, a human is capable of producing more than 1 trillion different antibody molecules. Millions of genes would be required to store the genetic information used to produce these receptors, but, the entire human genome contains fewer than 25,000 genes.
This myriad of receptors are produced through a process known as clonal selection.According to the clonal selection theory, at birth, an animal will randomly generate a vast diversity of lymphocytes (each bearing a unique antigen receptor) from information encoded in a small family of genes. In order to generate each unique antigen receptor, these genes will have undergone a process called V(D)J recombination, or combinatorial diversification, in which one gene segment recombines with other gene segments to form a single unique gene.
It is this assembly process that generates the enormous diversity of receptors and antibodies, before the body ever encounters antigens, and enables the immune system to respond to an almost unlimited diversity of antigens.Throughout the lifetime of an animal, those lymphocytes that can react against the antigens an animal actually encounters, will be selected for action, directed against anything that expresses that antigen.
It is important to note that the innate and adaptive portions of the immune system work together and not in spite of each other. The adaptive arm, B and T cells, would be unable to function without the input of the innate system. T cells are useless without antigen-presenting cells to activate them, and B cells are crippled without T-cell help. On the other hand, the innate system would likely be overrun with pathogens without the specialized action of the adaptive immune response.
Adaptive immunity during pregnancy
The cornerstone of the immune system is the recognition of "self" versus "non-self". Therefore, the mechanisms which protect the human fetus (which is considered "non-self") from attack by the immune system, are particularly interesting. Although no comprehensive explanation has emerged to explain this mysterious, and often repeated, lack of rejection, two classical reasons may explain how the fetus is tolerated. The first is that the fetus occupies a portion of the body protected by a non-immunological barrier, the uterus, which the immune system does not routinely patrol.The second is that the fetus itself may promote local immunosuppression in the mother, perhaps by a process of active nutrient depletion. A more modern explanation for this induction of tolerance is that specific glycoproteins expressed in the uterus during pregnancy suppress the uterine immune response (see eu-FEDS).

During pregnancy in viviparous mammals (all mammals except Monotremes), endogenous retroviruses are activated and produced in high quantities during the implantation of the embryo. They are currently known to possess immunosuppressive properties, suggesting a role in protecting the embryo from its mother's immune system. Also viral fusion proteins apparently cause the formation of the placental syncytium in order to limit the exchange of migratory cells between the developing embryo and the body of the mother (something an epithelium will not do sufficiently, as certain blood cells are specialized to be able to insert themselves between adjacent epithelial cells). The immunodepressive action was the initial normal behavior of the virus, similar to HIV, the fusion proteins were a way to spread the infection to other cells by simply merging them with the infected one (HIV does this too). It is believed that the ancestors of modern viviparous mammals evolved after an infection by this virus, enabling the fetus to survive the immune system of the mother.
The human genome project found several thousand ERVs classified into 24 families.
Hello Dude,
ReplyDeleteImmunity that is not inherited. Acquired immunity can be active or passive. Active immunity results from the development of antibodies in response to an antigen, as from exposure to an infectious disease or through vaccination. Thanks a lot!
Contract Research
Hello,
ReplyDeleteThanks for providing these useful tips over here. Adaptive immune system is made of specialized cells and processes which kill pathogens or prevent their attack, which is switched on by the evolutionarily older innate immune system...
Drug Discovery
https://bayanlarsitesi.com/
ReplyDeleteEskişehir
Erzincan
Ardahan
Erzurum
BXZ4K
Eskişehir
ReplyDeleteAdana
Sivas
Kayseri
Samsun
5UQS
Kocaeli
ReplyDeleteDenizli
Bursa
istanbul
Van
CH7
whatsapp görüntülü show
ReplyDeleteücretli.show
5TGK5M
Maraş Lojistik
ReplyDeleteHatay Lojistik
Tokat Lojistik
Elazığ Lojistik
Aksaray Lojistik
3E1Yİ3
43879
ReplyDeleteNiğde Parça Eşya Taşıma
Ankara Lojistik
Edirne Parça Eşya Taşıma
Çankırı Lojistik
Zonguldak Parça Eşya Taşıma
8F484
ReplyDeleteSincan Boya Ustası
Hatay Parça Eşya Taşıma
Silivri Boya Ustası
Muğla Parça Eşya Taşıma
Ordu Lojistik
Sakarya Şehir İçi Nakliyat
Ünye Çelik Kapı
Ünye Asma Tavan
Yalova Parça Eşya Taşıma
F4A47
ReplyDeletewinstrol stanozolol for sale
order halotestin
order primobolan
pharmacy steroids
Şırnak Evden Eve Nakliyat
Urfa Evden Eve Nakliyat
Balıkesir Evden Eve Nakliyat
sustanon
peptides for sale
626A3
ReplyDeleteÇerkezköy Çelik Kapı
Ardahan Parça Eşya Taşıma
Çerkezköy Ekspertiz
Hakkari Evden Eve Nakliyat
Adana Parça Eşya Taşıma
Osmaniye Şehir İçi Nakliyat
Tunceli Evden Eve Nakliyat
Bartın Parça Eşya Taşıma
Karabük Lojistik
0CBCE
ReplyDeletereferanskodunedir.com.tr
97749
ReplyDeleteaksaray görüntülü sohbet siteleri
mobil sohbet siteleri
bayburt telefonda kadınlarla sohbet
görüntülü sohbet siteleri
Çorum Parasız Görüntülü Sohbet Uygulamaları
Şırnak Sohbet Muhabbet
şırnak görüntülü canlı sohbet
tokat görüntülü sohbet canlı
çanakkale mobil sohbet siteleri
A8845
ReplyDeletekilis kızlarla canlı sohbet
Muş Goruntulu Sohbet
sesli mobil sohbet
parasız görüntülü sohbet uygulamaları
sesli sohbet sitesi
urfa rastgele görüntülü sohbet
aksaray rastgele canlı sohbet
canlı sohbet bedava
hatay sohbet chat
1049A
ReplyDeleteParasız Görüntülü Sohbet
Telegram Görüntüleme Satın Al
Tiktok İzlenme Hilesi
Telegram Görüntüleme Hilesi
Madencilik Nedir
Coin Kazma Siteleri
Aion Coin Hangi Borsada
Yeni Çıkan Coin Nasıl Alınır
Qlc Coin Hangi Borsada
05F21
ReplyDeleteKwai Beğeni Hilesi
Twitter Takipçi Hilesi
Coin Nasıl Kazılır
Omlira Coin Hangi Borsada
Aptos Coin Hangi Borsada
Shibanomi Coin Hangi Borsada
Parasız Görüntülü Sohbet
Ön Satış Coin Nasıl Alınır
Parasız Görüntülü Sohbet
AFA1E
ReplyDeleteCoin Üretme Siteleri
Pitbull Coin Hangi Borsada
Coin Kazanma
Binance Referans Kodu
Binance Referans Kodu
Omlira Coin Hangi Borsada
Coin Üretme
Madencilik Nedir
Soundcloud Takipçi Hilesi
52DD7
ReplyDeleteledger desktop
quickswap
dextools
uniswap
bitbox
onekey
sushi
arbitrum
layerzero
AF88B
ReplyDeleteİliç
Ağlasun
Bünyan
SAMSUN
Sütçüler
Düziçi
Yunak
Borçka
Ladik
HGMJVHJMK
ReplyDeleteتسليك مجاري بالقطيف
xbxcbnghbncgfjhn
ReplyDeleteرقم مصلحة المجاري بالاحساء
xcvbxcbxcbncvncvb
ReplyDeleteرقم مصلحة المجاري بالاحساء
شركة كشف تسربات المياه بالدمام 9is7c9rfI0
ReplyDeleteشركة عزل اسطح بخميس مشيط AjcQm93IDZ
ReplyDeleteشركة مكافحة الحمام بالاحساء oSXrFAe2s0
ReplyDeleteرقم المجاري بالاحساء AWa8wggJqu
ReplyDeleteشركة مكافحة النمل الابيض بابها M6NXzksAhb
ReplyDelete8DD060144C
ReplyDeleteşov
www.ijuntaxmedikal.store
görüntülü show
cialis
steroid satın al
steroid satın al
شركة عزل اسطح بخميس مشيط m7FcoDzn5Q
ReplyDeleteشركة تنظيف مدارس بجازان 7AwnEiUtmR
ReplyDelete23FE0EFDD1
ReplyDeleteAnadoluslot
Anadoluslot
Anadoluslot Güncel Giriş Adresi
Anadoluslot Yeni Adres
Trwin Güncel Giriş Adresi
Trwin
Trwin Yeni Adres
Trwin
Trwin
B1D3B81CA8
ReplyDeleteinstagram çok ucuz takipçi
instagram beğeni satın al
gerçek takipçi
ig takipçi
mobil ödeme takipçi
A2B74BA2D8
ReplyDeletetwitter bot takipçi
begeni satin al
bayan takipçi
takipçi
garantili takipçi