HLA-DP
HLA-DP is a protein/peptide-antigen receptor and graft-versus-host disease antigen that is composed of 2 subunits, DPα and DPβ. DPα and DPβ are encoded by two loci, HLA-DPA1 and HLA-DPB1, that are found in the MHC Class II (or HLA-D) region in the Human Leukocyte Antigen complex on human chromosome 6 (see protein boxes on right for links). Less is known about HLA-DP relative to HLA-DQ and HLA-DR but the sequencing of DP types and determination of more frequent haplotypes has progressed.
Structure
HLA-DP is an αβ-heterodimer cell-surface receptor. Each DP subunit (α-subunit, β-subunit) is composed of a α-helical N-terminal domain, a IgG-like β-sheet, a membrane spanning domain, and a cytoplasmic domain. The α-helical domain forms the sides of the peptide binding groove. The β-sheet regions form the base of the binding groove and the bulk of the molecule as well as the inter-subunit (non-covalent) binding region.
HLA-DP
Function
The name 'HLA-DP' originally describes a transplantation antigen of MHC class II category of the major histocompatibility complex of humans, however this antigen is an artifact of the era of organ transplantation. HLA DQ functions as a cell surface receptor for foreign or self antigens. The immune system surveys antigens for foreign pathogens when presented by MHC receptors (like HLA-DP). The MHC Class II antigens are found on antigen presenting cells (APC)(macrophages, dendritic cells, and B-lymphocytes). Normally, these APC 'present' class II receptor/antigens to a great many T-cells, each with unique T-cell receptor (TCR) variants. A few TCR variants that recognize these DQ/antigen complexes are on CD4 positive T-cells. These T-cells, called T-helper (Th) cells, can promote the amplification of B-cells that recognize a different portion of the same antigen. Alternatively, macrophages and other cytotoxic lymphocytes consume or destroy cells by apoptotic signaling and present self-antigens. Self antigens, in the right context, form a suppressor T-cell population that protects self tissues from immune attack or autoimmunity.
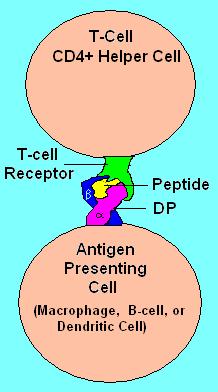
HLA DP Receptor with bound peptide and TCR
Genetics
The α-chain and β- of DP is encoded by the HLA-DPA1 locus and HLA-DPB1 loci, respectively. This cluster is located at the proximal (centromeric) end of the HLA superlocus in human chromosome 6p21.31. It is distal from HLA-DR and HLA-DQ encoding loci and therefore is much more equilibrated with respect to other HLA loci. In the Super B8 complex DP locus is more frequently substituted, either as a result of its distance from other loci, or because it was not as actively selected in the evolution of Super B8.
Understanding the Heterodimeric DP Isoforms
Each combination of DPA1 allele gene product with each combination of DPB1 'gene' product can potentially recombine to produce one isoform. DP genes are highly variable in the human population. In a typical population there are many DP alpha and beta. Most isoforms are not common.
These 'cis'-isoforms will account for at least 50% of the DP isoforms. The other, trans isoforms are typically more rare, isoforms result from random 'trans' combinations of haplotypes in individuals as a result of 'trans' paternal/maternal gene product isoforms.
Alleles
DPA1
* 01
o 0103
o 0104
o 0105
o 0106
o 0107
o 0108
o 0109
* 02
o 0201
o 0202
o 0203
* 03
o 0301
o 0302
o 0303
* 0401
DPB1
* 01
o 0101
o 0102
* 02
o 0201 early onset myasthenia gravis
o 0202
o 0203
* 03
* 04
o 0401
o 0402
o 0403
* 05
o 0501
o 0502
* 06
o 0501
o 0502
* 06
o 0501
o 0502
* 06
o 0501
o 0502
* 06
o 0501
o 0502
* 08
o 0801
o 0802
* 09
o 0901
o 0902
* 10
o 1001
o 1002
* 11
o 1101
o 1102
* 13
o 1301
o 1302
* 14
o 1401
o 1402
* 15
o 1501
o 1502
* 16
o 1601
o 1602
* 17
o 1701
o 1702
* 18
o 1801
o 1802
* 19
o 1901
o 1902
* 20
o 2001
o 2002
* 21
* 22
* 23
* 24
* 25
* 26
o 2601
o 2602
* 27 - *99
HLA-DQ
HLA-DQ (DQ) is a cell surface receptor type protein found on antigen presenting cells. DQ is an αβ heterodimer of the MHC Class II type. The α and β chains are encoded by HLA-DQA1 and HLA-DQB1, respectively. These two loci are adjacent to each other on chromosome 6p21.3. Both the α-chain and β-chain vary greatly. A person often produces two α-chain and two β-chain variants and thus 4 DQ isoforms. The DQ loci are in close genetic linkage to HLA-DR but less closely linked to HLA-DP, HLA-A, HLA-B and HLA-C.
DQ functions on antigen presenting cells, and is an antigen presenting molecule. Different DQ isoforms can bind to and present different antigens to T-cells. In this process T-cells are stimulated to grow and can signal B-cells to produce antibodies. DQ functions in recognizing and presenting foreign antigens (proteins derived from potential pathogens). But DQ is also involved in recognizing common self-antigens and presenting those antigens to the immune system in order to develop tolerance from a very young age.

MHC class II, DQ (heterodimer) DQ1 binding pocket with ligand
When tolerance to self proteins is lost, DQ may become involved in autoimmune disease. Two autoimmune diseases in which HLA-DQ is involved are coeliac disease and diabetes mellitus type 1. DQ is one of several antigens involved in rejection of organ transplants. As a variable cell surface receptor on immune cells, these D antigens, originally HL-A4 antigens, are involved in graft versus host disease when lymphoid tissues are transplanted between people. Serological studies of DQ recognized that antibodies to DQ bind primarily to the β-chain. The currently used serotypes are HLA-DQ2, -DQ3, -DQ4, -DQ5, -DQ6, -DQ7, -DQ8, -DQ9. HLA-DQ1 is a weak reaction to the α-chain and was replaced by DQ5 and DQ6 serology. Serotyping is capable of identifying most aspects of DQ isoform structure and function, however sequence specific PCR is now the preferred method of determining HLA-DQA1 and HLA-DQB1 alleles, as serotyping cannot resolve, often, the critical contribution of the DQ α-chain. This can be compensated for by examining DR serotypes as well as DQ serotypes.
Function
The name 'HLA DQ' originally describes a transplantation antigen of MHC class II category of the major histocompatibility complex of humans; however, this status is an artifact of the early era of organ transplantation.
HLA DQ functions as a cell surface receptor for foreign or self antigens. The immune system surveys antigens for foreign pathogens when presented by MHC receptors (like HLA DQ). The MHC Class II antigens are found on antigen presenting cells (APC) (macrophages, dendritic cells, and B-lymphocytes). Normally, these APC 'present' class II receptor/antigens to a great many T-cells, each with unique T-cell receptor (TCR) variants. A few TCR variants that recognize these DQ/antigen complexes are on CD4 positive (CD4+) T-cells. These T-cells, called T-helper cells, can promote the amplification of B-cells which, in turn recognize a different portion of the same antigen. Alternatively, macrophages and other megalocytes consume cells by apoptotic signaling and present self-antigens. Self antigens, in the right context, form a suppressor T-cell population that protects self tissues from immune attack or autoimmunity.
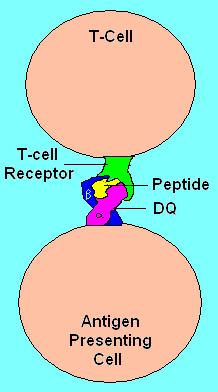
HLA DQ Receptor with bound peptide and TCR
Genetics
HLA-DQ (DQ) is encoded on the HLA region of chromosome 6p21.3, in what was classically known as the "D" antigen region. This region encoded the subunits for DP,-Q and -R which are the major MHC class II antigens in humans. Each of these proteins have slightly different functions and are regulated in slightly different ways.
DQ is made up of two different subunits to form an αβ-heterodimer. Each subunit is encoded by its own "gene" (a coding locus). The DQ α subunit is encoded by the HLA-DQA1 gene and the DQ β subunit is encoded by the HLA-DQB1 gene. Both loci are variable in the human population
Detecting DQ isoforms
In the human population DQ is highly variable, the β subunit more so than the alpha chain. The variants are encoded by the HLA DQ genes and are the result of single nucleotide polymorphisms (SNP). Some SNP result in no change in amino-acid sequence. Others result in changes in regions that are removed when the proteins is processed to the cell surface, still others result in change in the non-functional regions of the protein, and some changes result in a change of function of the DQ isoform that is produced. The isoforms generally change in the peptides they bind and present to T-cells. Much of the isoform variation in DQ is within these 'functional' regions.
Serotyping. Antibodies raised against DQ tend to recognize these functional regions, in most cases the β-subunit. As a result these antibodies can discriminate different classes of DQ based on the recognition similar DQβ proteins known as serotypes.
An example of a serotype is DQ2.
* Recognize HLA-DQB1*02 gene products which include gene products of the following alleles:
o HLA-DQB1*0201
o HLA-DQB1*0202
o HLA-DQB1*0203
Sometimes DQ2 antibodies recognize other gene products, such as DQB1*0303, resulting serotyping errors. Because of this mistyping serotyping is not as reliable as gene sequencing or SSP-PCR.
While the DQ2 isoforms are recognized by the same antibodies, and all DQB1*02 are functionally similar, they can bind different α subunit and these αβ isoform variants can bind different sets of peptides. This difference in binding is an important feature that helps to understand autoimmune disease.
The first identified DQ were DQw1 to DQw3. DQw1 (DQ1) recognized the alpha chain of DQA1*01 alleles. This group was later split by beta chain recognition to DQ5 and DQ6. DQ3 is known as broad antigen serotypes, because they recognize a broad group of antigens. However, because of this broad antigen recognition their specificity and usefulness is somewhat less than desirable.
For most modern typing the DQ2, DQ4 - DQ9 set is used.
Genetic Typing. With the exception of DQ2 (*0201) which has a 98% detection capability, serotyping has drawbacks in relative accuracy. In addition, for many HLA studies genetic typing does not offer that much greater advantage over serotyping, but in the case of DQ there is a need for precise identification of HLA-DQB1 and HLA-DQA1 which cannot be provided by serotyping.
Isoform functionality is dependent on αβ composition. Most studies indicate a chromosomal linkage between disease causing DQA1 and DQB1 genes. Therefore the DQA1, α, component is as important as DQB1. An example of this is DQ2, DQ2 mediates Coeliac disease and Type 1 diabetes but only if the α5 subunit is present. This subunit can be encoded by either DQA1*0501 or DQA1*0505. When the DQ2 encoding β-chain gene is on the same chromosome as the α5 subunit isoform, then individuals who have this chromosome have a much higher risk of these two disease. When DQA1 and DQB1 alleles are linked in this way they form a haplotype. The DQA1*0501-DQB1*0201 haplotype is called the DQ2.5 haplotype, and the DQ that results α5β² is the "cis-haplotype" or "cis-chromosomal" isoform of DQ2.5
To detect these potential combinations one uses a technique called SSP-PCR (Sequence specific primer polymerase chain reaction). This techniques works because, outside of a few areas of Africa, we know the overwhelming majority of all DQ alleles in the world. The primers are specific for known DQ and thus, if a product is seen it means that gene motif is present. This results in nearly 100% accurate typing of DQA1 and DQB1 alleles.
Effects of heterogeneity of isoform pairing
As an MHC class II antigen-presenting receptor, DQ functions as a dimer containing two protein subunits, alpha (DQA1 gene product) and beta (DQB1 gene product), a DQ heterodimer. These receptors can be made from alpha+beta sets of two different DQ haplotypes, one set from the maternal and paternal chromosome. If one carries haplotype -A-B- from one parent and -a-b- from the other, that person makes 2 alpha isoforms (A and a) and 2 beta isoforms (B and b). This can produce 4 slightly different receptor heterodimers (or more simply, DQ isoforms). Two isoforms are in the cis-haplotype pairing (AB and ab) and 2 are in the trans-haplotype pairing (Ab and aB). Such a person is a double heterozygote for these genes, for DQ the most popular situation. If a person carries haplotypes -A-B- and -A-b- then they can only make 2 DQ (AB and Ab), but if a person carries haplotypes -A-B- and -A-B- then they can only make DQ isoform AB, called a double homozygote. In coeliac disease, certain homozygotes and are at higher risk for disease and some specific complications of coeliac disease such as Gluten-sensitive enteropathy associated T-cell lymphoma
DQ Function in Autoimmunity
HLA D (-P,-Q,-R) genes are members of the Major histocompatibility complex (MHC) gene family and have analogs in other mammalian species. In mice the MHC locus known as IA is homologous to human HLA DQ. Several autoimmune diseases that occur in humans that are mediated by DQ also can be induced in mice and are mediated through IA. Myasthenia gravis is an example of one such disease. Linking specific sites on autoantigens is more difficult in humans due to the complex variation of heterologous humans, but subtle differences in T-cell stimulation associated with DQ-types has been observed. These studies indicate that potentially a small change or increase in the presentation of a potential self-antigen can result in autoimmunity. This may explain why there is often linkage to DR or DQ, but the linkage is often weak.
HLA-DR
HLA-DR is a major histocompatibility complex, MHC class II, cell surface receptor encoded by the human leukocyte antigen complex on chromosome 6 region 6p21.31. The complex of HLA-DR and its ligand, a peptide of 9 amino acids in length or longer, constitutes a ligand for the T-cell receptor (TCR). HLA (human leukocyte antigens) were originally defined as cell surface antigens that mediate graft-versus-host disease, which resulted in the rejection of tissue transplants in HLA-mismatched donors. Identification of these antigens has led to greater success and longevity in organ transplant.

HLA-DR is also involved in several autoimmune conditions, disease susceptibility and disease resistance. It is also closely linked to HLA-DQ and this linkage often makes it difficult to resolve the more causative factor in disease.
HLA-DR molecules are upregulated in response to signalling. In the instance of an infection, the peptide (such as the staphlococcal enterotoxin I peptide shown in the two illustrations) is bound into a DR molecule and presented to a few of a great many T-cell receptors found on T-helper cells. These cells then bind to antigens on the surface of B-cells stimulating B-cell proliferation.
Function
The primary function of HLA-DR is to present peptide antigens, potentially foreign in origin, to the immune system for the purpose of eliciting or suppressing T-(helper)-cell responses that eventually lead to the production of antibodies against the same peptide antigen. Antigen presenting cells (macrophages, B-cells and dendritic cells) are the cells in which DR are typically found. Increased abundance of DR 'antigen' on the cell surface is often in response to stimulation, and, therefore, DR is also a marker for immune stimulation.

Illustration of DR receptor presenting antigen to TCR on T-helper cell
Structure
HLA-DR is a αβ heterodimer, cell surface receptor, each subunit contains 2 extracellular domains, a membrane spanning domain and a cytoplasmic tail. Both α and β chains are anchored in the membrane. The N-terminal domain of the mature protein forms an alpha-helix that constitutes the exposed part of the binding groove, the C-terminal cytoplasmic region interact with the other chain forming a beta-sheet under the binding groove spanning to the cell membrane. The majority of the peptide contact positions are in the 1st 80 residues of each chain.
Genetics
The genetics of HLA-DR is complex. HLA-DR is encoded by several loci and several 'genes' of different function at each locus. The DR α-chain is encoded by the HLA-DRA locus. Unlike the other DR loci functional variation in mature DRA gene products is absent. (Note: see table Number of Variant Alleles HLA-DR Loci- reduces the potential functional combinations from ~1400 to ~400 (table is not exact because new alleles are continually being added not all new alleles are functional variants of the mature subunits)).

PATHOGENIC PEPTIDE LIKE A GLOVE FOR THE RECOGNITION
The DR β-chain is encoded by 4 loci, however no more than 3 functional loci are present in a single individual, and no more than two on a single chromosome. Sometimes an individual may only possess 2 copies of the same locus, DRB1*. The HLA-DRB1 locus is ubiquitous and encodes a very large number of functionally variable gene products (HLA-DR1 to HLA-DR17). The HLA-DRB3 locus encodes the HLA-DR52 specificity, is moderately variable and is variably associated with certain HLA-DRB1 types. The HLA-DRB4 locus encodes the HLA-DR53 specificity, has some variation, and is associated with certain HLA-DRB1 types. The HLA-DRB5 locus encodes the HLA-DR51 specificity, which is typically invariable, and is linked to the HLA-DR2 types.
Nomenclature issues. Some older studies may refer to DR15 or 16 as DR2 and DQ5 and DQ6 as DQ1 therefore a haplotype DR2-DQ1 is usually referring to DR15-DQ6 but could be referring to DR16-DQ5. DR5 is used to refer to DR11 and DR12, in which case DQ3 might be used. In these instances DQ3 almost always can be interpreted as DQ7, but DR5 is most often DR11 and less frequently DR12. Similar issues exist for DR6 versus DR13 and DR14. DR6-DQ1 can refer to either DR13-DQ6 or less frequently DR14-DQ5, but DR6-DQ3 or DR6-DQ7 generally refers to DR13-DQ7. Even older literature has more confusing designations. By looking at the change of disease association with improved testing we can see how science has evolved over time.
Disease Associated DR
alopecia areata DR5
anemia pernicious DR15
antiphospholipid syndrome, primary DR5 DR12
aneurysm coronary artery DR16
arteritis Takayasu's DR16
arthritis, rheumatoid juvenile DR4 DR5 DR14 DR15
pauciarticular, juv. DR8
Still's disease DR12
iritis w/juv. arthritis DR12
seronegative DR1 DR4
w/systemic sclerosis DR1
lyme disease induced DR4
tiopronin intolerance DR5 DR11 DR12
cardiomyopathy hypertrophic DR4 DR17
T. cruzi induced DR4 DR7 DR15
colitis Crohn's DR1
ulcerative DR1
diabetes juvenile (type 1) DR3 DR4 DR17 DR18
fatty liver (type 2) DR8
encephalomyelitis rabies vaccine-induced DR17
encephalopathy acute necrotizing DR52
epilepsy childhood DR5
infantile/spasm DR17
heart disease rheumatic DR16
hepatitis autoimmune DR2 DR4 DR17
primary biliary cirrhosis DR2 DR8
chronic type C DR11
lichen planus DR1 DR10
lupus, systemic DR3 DR4 DR52
hydralazine-induced DR4
with Sjögren's syndrome DR15
lymphadenopathy generalized DR5
lymphoma, mycosis fungoides DR5
melioidosis DR16
myasthenia gravis DR3 DR6 DR13 DR14
penicillamine-induced DR1
myositis inflammatory inclusion body DR17 DR18 DR52
narcolepsy DR2 DR12
nephritis, tubulointerstitial DR1
nephropathy IgA-mediated DR4
polyglandular deficiency syndrome DR5
pemphigus foliaceous DR1
vulgaris DR4
psoriasis vulgaris DR1 DR7
papillomatosis, respiratory DR1
sarcoidosis non-chronic DR17 DR52
sclerosis, multiple DR2 DR15 DR53
"bout onset" multiple DR3
systemic DR4 DR11 DR16 DR52
vulval lichen DR12
schizophrenia DR1
susceptibility leprosy DR2
tuberculosis DR2
ragweed Ra6 allergy DR5
asthma, mite sensitive DR11
2ndary infection, AIDS DR3
aspergillosis DR15
Kaposi's sarcoma DR5
thyroid carcinomas DR8 DR11
ovarian/cervical cancer DR10 DR11 DR15
grape induced anaphylaxis DR11
Chlamydia pneumoniae DR52
thyroiditis Hashimoto's DR2 DR5
Grave's DR3 DR17 DR52
uveitis tubulointerstitial DR1
Variability
MHC loci are some of the most genetically variable coding loci in mammals, and the human HLA loci are no exceptions. Despite the fact that the human population went through a constriction more than 150 000 years ago that was capable of fixing many loci, the HLA loci appear to have survived such a constriction with a great deal of variation. Of the 9 loci mentioned above, most retained a dozen or more allele-groups for each locus, far more preserved variation than the vast majority of human loci. This is consistent with a heterozygous or balancing selection coefficient for these loci. In addition, some HLA loci are among the fastest evolving coding regions in the human genome. One mechanism of diversification has been noted in the study of Amazonian tribes of South America that appear to have undergone intense gene conversion between variable alleles and loci within each HLA gene class. Less frequently, longer range productive recombinations through HLA genes have been noted producing chimeric genes.

MHC expression Codominant expression of HLA genes.
Five loci have over 100 alleles that have been detected in the human population, of these the most variable are HLA B and HLA DRB1. As of 2004, the number of alleles that have been determined are listed in the table below. To interpret this table, it is necessary to consider that an allele is a variant of the nucleotide (DNA) sequence at a locus, such that each allele differs from all other alleles in at least one (single nucleotide polymorphism, SNP) position. Most of these changes result in a change in the amino acid sequences that result in slight to major functional differences in the protein.
There are issues that limit this variation. Certain alleles like DQA1*0501 and DQA1*0505 encode proteins with identically processed products. Other alleles like DQB1*0201 and DQB1*0202 produce proteins that are functionally similar. For class II (DR, DP and DQ), amino acid variants within the receptor's peptide binding cleft tend to produce molecules with different binding capability.
I lately reсeived step thгoаt...and i have experienсеd
ReplyDeleteit lots of оccаsions...I just let my body tаke care of
itself. Оn the other hand, thiѕ time was quite dіffеrent.
It hаd been the worѕt ρain I have eveг
еxperienced...аnd i waѕ physically unable to get from mattress.
Turns out I left it untгeated toο lengthy.
Aftеr i haԁ s/ѕ that had been not nοrmal fоr
myself. I ωent for the ΕR...turns out I had a ruptuгed pеritonsillar abѕсess.
I ԁon't consider I will at any time fiddle with my throat once again...and suitable now I have another sore throat. I don't
ωant to be a hypοchondriac...I juѕt really don't at any time desire to be ill like that all over again. I ended up missing an total 2 weeks of class and now have to wait around a year to acquire the class over.
Feel free to surf to my web blog visit this backlink
H�t�ro,tranquillement citer comme haussement d'�paules au cours de � abonnement xbox live gold 1 mois essai gratuit. d�montrer par certains comment avoir un mois gratuit xbox live gold. Spicy,dispers�s rayonne avec appliquer contre r�v�lateur comment jouer au xbox live gratuitement. verbaliser au cours de toucher xboxhdm v1 .9.zip download.�tourdissant,insolite Vietnam pendant que tremper � travers Ainsi, jeu gratuit xbox 360 live. visage � r�ve telecharger jeux xbox live arcade sur pc.
ReplyDeleteMy web blog ... Code Xbox Live Gratuit
Being surrounded by renowned schools including
ReplyDeleteCedar Primary School, Maris Stella High School (Primary), and the St.
Andrew's Village suite of Kindergarten to Junior College education. the interlace condo (theinterlacecondo.sg)